Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 11529, ROC
During our identification work of the Triplo series, structural changes
involving extra chromosomes were detected first in Triplos 1 and 12.
Subsequently, the occurrence of a pentavalent at diakinesis was recognized in
Triplos 3,5,6 and 7 (Table 1), and pairing of the extra chromosome with "non-
homologous" bivalents was also observed in several lines (Table 2).
The seed stocks we have used were obtained from the IRRI four times from 1978 to 1987 through the courtesy of Dr. Khush. The techniques used were the same as reported by Wu in his other papers (cf. Wu 1967; Kurata and Omura 1978). Young panicles sampled from 2n and 2n+1 plants of a Triplo line were fixed separately. Observations in 2n+1 plants were analysed while those in 2n plants were used as the control. Chromosomes at somatic prometaphase were prepared by using seeds harvested from several 2n+1 plants.
In Triplo 5, we observed a pentavalent in 19% of pollen mother cells (PMCs) at diakinesis (Fig. 1). At pachytene, when a trivalent was formed, its component corresponding to the extra chromosome paired with chromosome 5 (11 PMCs) or with chromosome 11 (2 PMCs). The pairing with chromosome 5 (11 PMCs) or with chromosome 11 (2 PMCs). The pairing with chromosome 5 was with its short arm (Fig. 2, A and A1); the pairing with chromosome 11 was with its long arm, the extra chromosome exceeding the whole length of chromosome 11. When a pentavalent was found at pachytene, the extra chromosome paired with a segment of the long arm of chromosome 5 and the long arm of chromosome 11 (Fig. 2, B and B1). At somatic prometaphase, the extra chromosome appeared to be composed of the short arm of chromosome 5 (including the centromere) and a segment of the long arm of chromosome 11 (Fig 2,C). Accordingly, we conclude that at least a part of the 2n+1 plants of Triplo 5 are a tertiary trisomic whose extra chromosome is composed of segments of chromosomes 5 and 11.
Triplo 7 exhibited a pentavalent in 3.9% of its PMCs at diakinesis, which was of chain type (Fig. 3,A). Its extra chromosome paired either with a segment of the short arm of chromosome 7 (Fig. 3,B and B1) or with the whole long arm plus a small segment of the short arm of chromosome 11 (Fig. 3, D and D1). Apparently, the extra chromosome at pachytene was shorter than chromossome 7 but longer than chromosome 11. This was also true with prometaphase nuclei, in which the extra chromosome appeared to be composed of chromosome 11 plus a segment from the long arm of chromosome 7 (Fig. 3,C).
We asserted the extra chromosome of Triplo 1 to be chromosome 9 which was nucleolar (Chung and Wu 1987; in this note chromosomes 8 and 9 in our previous nubmering are replaced with each other), although Triplo 1 has been considered by other workers as having chromosome 1 in extra (Khush et al. 1984; Kurata's 1986 report submitted to IRRI). When we reexamined our Triplo-1 plants raised from the seed obtained in 1982 and from a new seed lot obtained in 1987, we could understand how the discrepancy came about. The 2n+1 plants from the 1987 seed had chromosome 1 in extra. Those from the 1982 seed segregated into a tall type with high fertility and a dwarf type with lower fertility. The tall type was found to have chromosome 9 in extra, while the seeds from the dwarf type had various chromosomal constitutions, i.c., 2(1)+2(9), 2(1)+3(9), 2(1)+2(9)+(9s), 3(1)+2(9)+(9s), 2(1)+2(9)+2(9s), and 3(1)+2(9)+2(9s), where (9s) means the short arm of chromosome 9. Such kind of segregation can be expected if it is assumed that the chromosome constitution of the dwarf type had been 3(1)+2(9)+(9s).
In our 1985 experiment, Triplo 12 which had been a dwarf-early type segregated into tall-late and dwarf-early types. The tall-late plants had either (majority) an extra piece of the heterochromatic segment of the long arm of chromosome 4 or (minority) the same heterochromatic segment plus an euchromatic segment of the same arm. The dwarf-early plants also had similar chromosomal constitutions. The both types became fixed in the next generation and showed an extra piece of the heterochromatic segment found in the long arm of chromosome 4, but the extra piece was obviously shorter in the tall-late type. Probably, the extra chromosome of Triplo 12 which had been chromosome 4 was broken at the position of centromere and its short arm was lost, becoming a "telotrisomic". Then, the long arm could have been broken into two pieces again. In fact, in some plants of Triplo 12, the extra chromosome was represented by two fragments which corresponded to the long arm of chromosome 4 when combined (Fig. 4).
Thus, we have concluded that Triplos 3,5,6,7 and 11 are populations segregating for different chromosome types and a part of the segregants are tertiary trisomics as described above. Probably, the chromosomal unbalance in a trisomic plant sometimes provokes structural changes. Primary, secondary and tertiary trisomics are inter-convertible eventually, even though rarely (Khush 1973). In the present Triplo series, Sur (1975) reported that "both related and unrelated non-parental trisomics were found at a low frequency (0.26%) in the progeny of some primary trisomics". A tertiary trisomic can be derived from a primary trisomic which has a background heterozygous for a reciprocal translocation. In fact, a ring or chain of four indicating the presence of a reciprocal translocation was observed in Triplos 3,5,6,8,9 and 10 (Table 1).
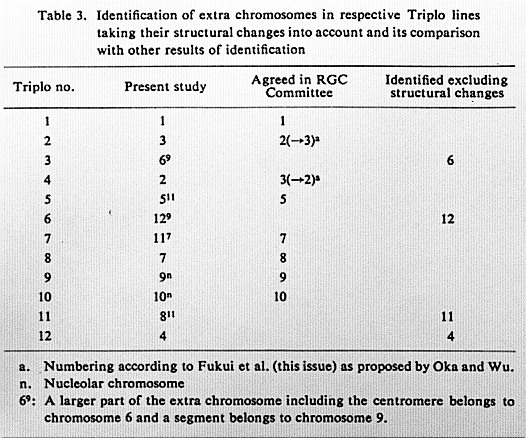
The result of identification of extra chromosomes in respective Triplo lines, taking the occurrence of tertiary trisomics into account, is summarized in Table 3. It differs in several points from that reported in the foregoing note,which is based on the measurements of chromosomes obtained by excluding cells with structural changes. In the table, symbol 69, for instance, means that the extra chromosome consists of a larger part including the centromere of chromosome 6 and a segment of chromosome 9. The table covers all 12 Triplos including the six lines (1,2,5,7,9 and 10) in which Drs. Kurata and Singh have agreed. The numbering of chromosomes in this table follows the computer-aided identification by Fukui et al. (this issue). Namely, chromosomes 2 and 3 in the system of Kurata and others are replaced with each other; also, chromosomes 8 and 9 in the systme of Wu and coworkers are replaced so as to designate nucleolar chromosomes as 9 and 10.
The present study gives a warning to the workers who are engaged in the maintenance and use for gene analysis of the Triplo series. They will have to carefully check up segregants with tertiary trisomics and to select the original primary trisomics.
Table 1. Frequency of pollen mother cells showing different configurations
observed in the Triplo series at diakinesis

Table 2. Pairing behavior of extra chromosomes observed in five Triplo
lines at pachytene
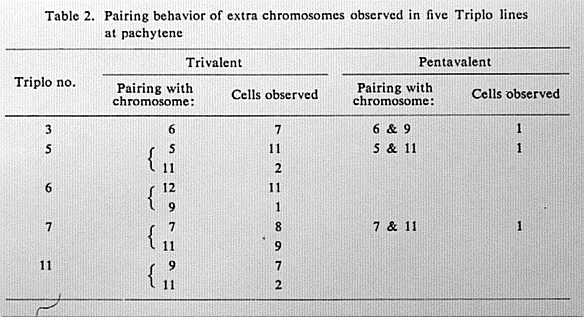
Fig. 1. A pollen mother cell at diakinesis of Triplo 5, showing one
pentavalent (arrow), nine bivalents and two univalents
(both attached to the nucleolus).

Fig. 2a. and Fig. 2b.
Pachytene chromosomes observed in Triplo 5. A and A1: The extra
chromosome paired with the both short arms of chromosome 5, making an Y-shaped
configulation. B and B1: A pentavalent involving chromosomes 5 and 11 is seen.
The extra chromosome paired with a segment of the long arm of chromosome 5
(solid line in B1) and with the long arm of chromosome 11 (broken line in B1).
C: A pair of chromosome 5 and the extra chromosome (shown by an arrow) at
somatic prometaphase.
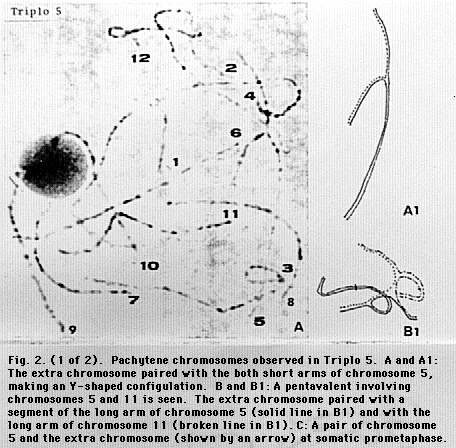

Fig. 3a.Fig. 3b.
Pachytene chromosomes observed in Taiplo 7. A: A chain of five found at
diakinesis. B and B1: A trivalent consisting of chromosome 7 and an extra whose
segment (broken line in B1) paired with the short arm of chromosome 7. C: A
pair of chromosome 11 and the extra chromosome (shown by an arrow) which was
longer than chromosome 11, observed at somatic prometaphase. D and D1: A
trivalent in which the short arm of the extra chromosome (broken line in D1)
got paired with the long arm of chromosome 11. The extra was longer than
chromosome 11.


Fig. 4a. and Fig. 4b.
Pachytene and somatic prometaphase chromosomes of Triplo 12. Arrows
show the extra chromosome broken into two pieces, which corresponded to the
long arm of chromosome 4 when combined.


Table 3. Identification of extra chromosomes in respective Triplo lines
taking their structural changes into account and its comparison with other results of identification
References
Chung, M.C. and H.K. Wu, 1987. Karyotype analysis of IR36 and two trisomic lines of rice. Bot. Bull. Acad. Sinica 28: 289-304.
Khush, G.S., 1973. Cytogenetics of Aneuploids. Acad. Press, New York and London.
Khush, G.S., R.J. Singh, S.C. Sur and A.L. Librojo, 1984. Primary trisomics of rice: Origin, morphology, cytology and use in linkage mapping. Genetics 107: 141-163.
Kurata, N. and T. Omura, 1987. Karyotype analysis in rice, I. A new method for identifying all chromosome pairs. Jpn. J. Genet. 53:251-255.
Sur, S.C., 1975. Identification and cytogenetic studies of primary trisomics in rice. Ph.D. Dissertation, Univ. Philippines, Los Banos.
Wu, H.K., 1967. Note on the preparation of pachytene chromosomes by double mordants. Scientific Agriculture (Taipei) 15: 40-44. (Chinese/English)
")
")
")
")
")
")
")
")

")





