activity in partially purified chloroplast soluble protein
fraction
National Agriculture Research Center, 3-1-1 Kannondai, Tsukuba,
Ibaraki, 305-8666 Japan
The differentiation of proplastids
into chloroplasts is accompanied by a drastic change in transcriptional
mechanisms. Particular photosynthesis genes encoded in the plastid genome
are specifically expressed during or after this conversion. Biochemical
studies have proved the existence of two types of RNA polymerase (RNAP)
in the chioroplast. One is nuclear encoded T7 bacteriophage type RNAP which
has been presumed to function in every type of plastid for transcribing
housekeeping genes like ribosomal components and tRNAs (Hedtke et a!. 1997).
Another is the chioroplast-specific enzyme sharing extensive similarities
with the bacterial a2BB'o RNAP. Genes corresponding to the bacterial counter
parts of the core enzyme, a2BB', have been found in the plastid genome
(Hiratsuka et al. 1989) and recently, genes encoding the a factor, which
is necessary for the transcriptional initiation, have been reported to
be encoded in the nuclear genomes of higher plants (Tanaka et a!. 1997,
Tozawa et al. 1998). The rice sigA gene shows shoot-specific and light-dependent
expression in seedlings and its amino-terminal region shows features of
potential plastid localization signals (Tozawa et a!. 1998). Transient
expression assay using GFP have revealed that amino-terminal region of
SigA homolog of Arabidopsis thaliana functions as a chloroplast targeting
signal (Isono eta!. 1998). Therefore, contribution of SigA to chloroplast-specific
gene expression has been proposed. To confirm its localization and correlation
with the activity of the bacterial type RNAP, immunoblot analysis and in
vitro transcription assays were performed.
Recombinant SigA was prepared using
the pQE vector kit (Qiagen) and rabbit antiserum against this protein was
prepared. Intact chioroplasts were isolated and purified from 10 day seedlings
of rice cv. Nipponbare, grown at 28°C under continuous light condition,
by percol gradient centrifugation (Orozco et at. 1985). Approximately 2
x i09 chloroplasts were obtained which were lysed in the high-salt buffer
containing 1M NaC1. They were then separated into two fractions: a soluble
fraction including mainly stromal proteins and an insoluble fraction including
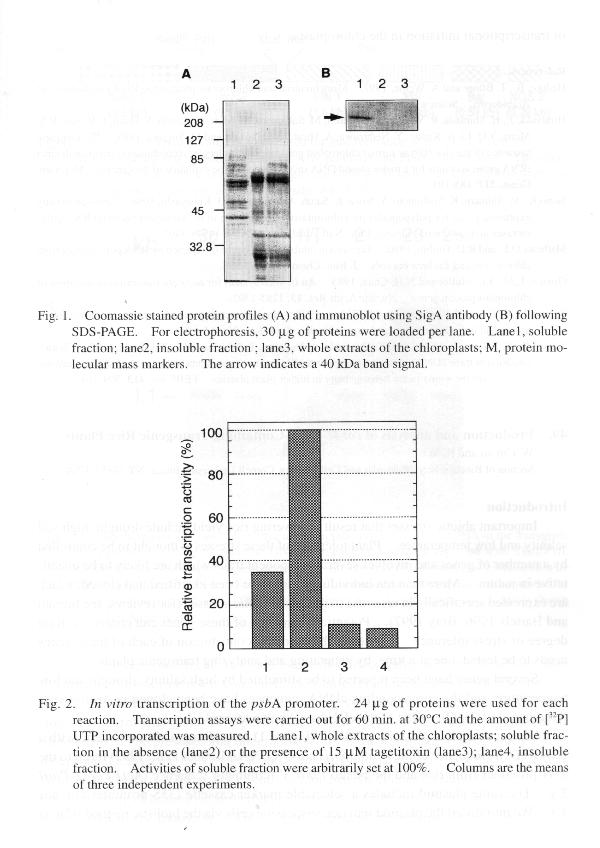
mainly proteins associated with thylakoid membranes. Polypeptide composition
pattern of those fractions are apparently different (Fig. 1A). For in vitro
transcription assay, -35/-10 type promoter of the rice psbA gene was used.
RNA polymerase activity was calculated based on the amount of 32P-labelled
UTP incorporated into RNA. Immunoblot analysis revealed that SigA is located
in the soluble fraction of the chloroplast (Fig. 1B), which is also associated
with the activity of bacterial RNAP (Fig. lB and 2). This transcriptional
activity was completely inhibited by supplemented tagetitoxin (Fig. 2),
the best known inhibitor of plastidial and bacterial RNAP (Mathews eta!.
1990). SigA protein conserves domains of promoter recognition, promoter
melting, and of core RNAP binding, those which have been assigned in bacterial
sigma factors (Tozawa eta!. 1998). Therefore, these observations suggest
crucial contribution of SigA, plastidial sigma factor coexisting with the
bacterial RNAP, to the process of transcriptional initiation in the chioroplasts.
Hedtke, B., T. Boner and A. Weihe, 1997. Mitochondrial and
chloroplast phage-type RNA polymerases in Arabidopsis. Science 277: 809-811.
Hiratsuka, J., H. Shimada, R. Whittier,T. Ishibashi, M. Sakamoto,
M. Mori, C. Kondo, Y. Honji, C.R. Sun, B.Y. Meng, Y.Q. Li, A. Kanno, Y.
Nishizawa, A. Hirai, K. Shinozaki and M. Sugiura, 1989. The complete sequence
of the rice (Oryza sativa) chloroplast genome: intermolecular recombination
between distinct tRNA genes accounts for a major plastid DNA inversion
during the evolution of the cereals. Mol. Gen. Genet. 217: 185-194.
Isono,K., M. Shimizu, K. Yoshimoto, Y. Niwa, K. Satoh, A.
Yokota and H. Kobayashi, 1998. Leaf-specifically expressed genes for polypeptides
for chloroplasts with domains of o70 factors of bacterial RNA polymerases
in Arabidopsis thaliana: Proc. Natl. Acad. Sci. USA 94: 14948-14953.
Mathews,D.E. and R.D. Durbin, 1990. Tagetitoxin inhibits
RNA synthesis directed by RNA polymerases from chloroplasts and Escherichia
coli. J. Biol. Chem. 265: 493-498.
Orozco, E.M., i.E. Mullet and N.H. Chua, 1985. An in vitro
system for accurate transcription initiation of chioroplast protein genes.
Nucleic Acids Res. 13: 1283-1302.
Tozawa,Y., K. Tanaka, H. Takahashi and K. Wakasa,l998. Nuclear
encoding of a plastid a factor in rice and its tissue- and light-dependent
expression. Nucleic Acids Res. 26: 415-419.
Tanaka,K., Y. Tozawa, N. Mochizuki, K. Shinozaki, A. Nagatani,
K. Wakasa and H. Takahashi, 1997. Characterization of three cDNA species
encoding plastid RNApolymerase sigma factors in Arabidopsis thaliana:
evidence for the sigma factor heterogeneity in higher plant
plastids. FEBS lett. 413: 309-313.
|
")
")
")
")
")
")
")
")

")





