progenies of rice
Plant Breeding Laboratory, Faculty of Agriculture, Kyushu
University, Fukuoka, 812-8581 Japan
Many genes controlling F1 hybrid sterility in intervarietal
crosses of Japonica and Indica have been reported and mapped. However,
little is known about the genes causing F2 sterility in such crosses. In
this study, a case of monogenic segregation of F2 sterility was found in
backcrossed F2 populations which were obtained during the development of
a series of substitution lines of Indica rice with a Japonica genetic background.
We report the chromosomal location of a gene controlling spikelet sterility
in F2 population.
To produce the substitution lines, recombinant inbred lines
derived from the cross between Japonica variety Asominori and Indica variety
1R24 (Tsunematsu et a!. 1996) were used for original crosses and backcrosses
with Asominori. Some of the BC3F2 plants derived by selfing BC3F1 plants
showed very low spikelet fertility (approximately 10% fertility). The difference
between fertile and sterile plants was clearly recognized. The segregation
of fertile and sterile plants agreed with a 3:1 ratio in some lines, suggesting
that the sterility was governed by a recessive gene. This recessive gene
is designated as s(t).
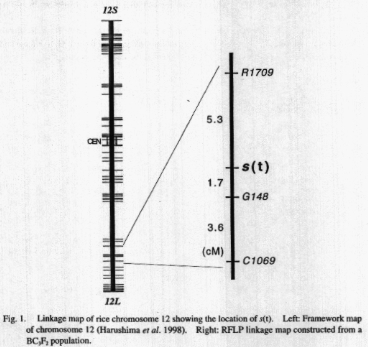
To determine the chromosomal location of s(t), introgressed
chromosomal region of BC3F1 plants segregating the sterile plants in their
selfed progenies was compared. Because the common region of chromosome
12 was introgressed in the BC3F1 plants, the gene controlling the spikelet
sterility was inferred to be on chromosome 12. Subsequently, several RFLP
markers in the putative region of chromosome 12 were used for RFLP mapping.
A BC3F2 population (n=57) which showed clear 3:1 segregation was used as
a mapping population. All the RFLP markers used agreed with 1:2:1 segregation
ratio in this BC3F2 population. Most of the sterile plants were homozygous
for 1R24 allele at the respective RFLP loci of chromosome 12 and close
linkage between s(t) and the RFLP loci was detected. The RFLP marker closest
to s(t) was G148 (Fig. 1).
Two genes Si5 and S17(t) for F1 sterility on chromosome
12 were previously identified (Wan et a!. 1996, Wan and Ikehashi 1995).
The relationship between these genes and s(t) in this study must be investigated.
From earlier works, it was assumed that F2 sterility was due to a set of
duplicate genes (Yokoo 1984). Since the F2 sterility in this study also
would be controlled by a set of duplicate genes, s(t) could be one of the
duplicate genes. We are now identifying the other gene(s).
This study was supported in part by the Program for Promotion
of Basic Research Activities for Innovative Biosciences.
|
|
References
Hanishima, Y., M. Yano, A. Shomura, M. Sato, T. Shimano,
Y. Kuboki, T. Yarnamoto, S.Y. Lin, B.A. Antonio, A.
Parco, H. Kajiya, N. Huang, K. Yamainoto, Y. Nagamura, N.
Kurata, G.S. Khush and T. Sasaki, 1998. A
high-density rice genetic linkage map with 2275 markers
using a single F2 population. Genetics 148:
479-494.
Tsunematsu, H., A. Yoshimura, Y. Harushima, Y. Nagamura,
N. Kurata, M. Yano, T. Sasaki and N. Iwata, 1996. RFLP framework map using
recombinant inbred lines in rice. Breed. Sci. 46: 279-284.
Yokoo, M., 1984. Female sterility in an indica-japonica
cross of rice. Jpn. J. Breed. 34: 219-227.
Wan, J. and H. Ikehashi, 1995. A new locus for hybrid sterility
in remote crosses of cultivated rice (Oryza saliva L.). VII. Breed. Sci.
45 (Suppl. 2): 191. (in Japanese)
Wan, J., Y. Yamaguchi, H. Kato and H. Ikehashi, 1996. Two
new loci for hybrid sterility in cultivated rice (Oryza saliva L.). Theor.
AppI. Genet. 92: 183-190.
|
")
")
")
")
")
")
")
")

")





