Use of tester lines is helpful in analyzing the expression
of heading time genes and nonallelic interaction between them. Among the
known heading time genes loci, Sel (Chandraratna 1955, Yano eta!. 1997)
and Efi (Tsai 1976), as well as El (Yamagata et a!. 1985), are especially
important because of their drastic action on photoperiod sensitivity and
basic vegetative growth period respectively. For the two loci, the following
three tester lines are useful because they have the same genotype for other
heading time loci (Okumoto et a!. 1992, Inoue et a!. 1998) : T65 (a variety
Taichung 65, SeleSele eflefi), T65Eb (an isogenic line of T65, Se1eSele
EflEfi), and EG7 (SelnSeln EflEfI). But neither any cultivars nor any lines
having the genotype SelnSeln eflefi have been found or produced as yet.
We planned to produce a new tester line having such a genotype
through the cross �EG7 x T65�. By selecting a few latest heading plants
in F2 to F11, a line fixed for a late heading time was obtained in the
F12. This line, coded EL3O, was crossed with T65 and T65Eb to identify
its genotype. The F2 populations were subjected to a gene analysis for
days to heading (heading time). The C locus was used as a linkage marker
of the Se) locus because these two loci are linked to each other on chromosome
6. T65 and T65Eb carry a dominant allele CB making apiculus purple in color
at ripening stage, while EL3O and its parental line EG7 carry a recessive
allele C+ not having such action.
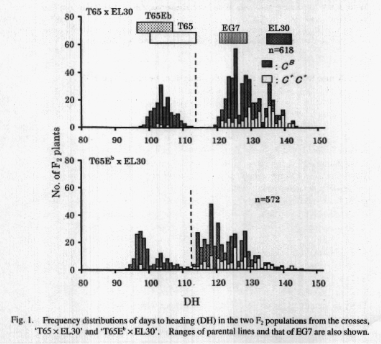
For days to heading, the F2 populations from crosses �T65
x EL3O� showed a bimodal distribution with a clear breakpoint between parental
ranges and without any distinct transgressive segregants (Fig.1). The breakpoint
divided the population into 163 early plants and 455 late plants, which
fitted to a 1 :3 ratio expected for one-locus segregation (x2=0.552, 0.50
> P> 0.25). Moreover, most C+C+ plants showed late heading. These results
indicate that T65 and EL3O differ only in the genotype for the Sel locus,
and eventually that both of them carry efi. The F2 population from the
cross �T65Eb x EL3O� showed a bimodal distribution without distinct transgressive
segregants, and a breakpoint divided the population into two groups: 163
early plants and 409 late plants, which fitted to a 1: 3 ratio expected
for one locus segregation (x2=3.730, 0.10> P> 0.50) (Fig. 1). Since most
C+C+ plants showed late heading, this segregation could be attributed to
the segregation of the Sel locus. Compared to the F2 population from the
cross �T65 x EL3O�, however, this population showed a wider range of distribution
in both groups, which was caused by the segregation of the Efi locus.
Line EL3O now shows fixation also for agronomic characters
other than heading time. The genotype of EL3O is shown in Table 1 along
with those of other tester lines used here. We are now conducting successive
backcrossing to raise isogenic tester lines of T65 for the Efi and Sel
loci.
|
|
Line
|
|
|
|
|
Efi
|
|
|
T65
|
|
eflefi
|
|
T65Eb
|
|
EflEfi
|
|
EG7
|
|
EflEfl
|
|
EL3O
|
|
eflefi
|
|
1)
|
All lines have proved to have the same genotype for other
heading time loci (Okumoto
|
")
")
")
")
")
")
")
")

")





