| RGN Home | Vol. 18 >C. Research Notes>III. Genetics of physiological traits and others |
| 28. | The effect of cytoplasmic male sterility factor in rice anther culture |
| S. M. BALACHANDRAN1, N. P. SARMA and E. A. SIDDIQ 1)Present Address: Tissue Culture & Genetic Engineering Lab, Plant Breeding, Genetics & Biochemistry Division International Rice Research Institute DAPO Box No. 7777, Metro Manila, The Philippines. |
| One of the key factors that determine anther culture response
is the genotype of the candidate crop species. Several studies have shown
that it is largely the nuclear genes that control in vitro response.
Although, the role of nuclear genomes in androgenic culture response has
been well established, the influence of specific cytoplasmic factor(s) such
as male sterility gene affecting anther/tissue culture has not been studied
to the desired extent. Understanding the cytoplasmic sterility effect on
anther culture response would require near-isogenic lines with and without
fertility restoration genes in identical sterilizing cytoplasm (Becraft
and Taylor 1989). We made an attempt to understand the effect of cytoplasmic
sterility factor in rice anther culture by comparing the anther culture
response of cytoplasmic male sterile (cms) line-based hybrids (A/R) with
their corresponding isonuclear-male fertile maintainer line-based hybrids
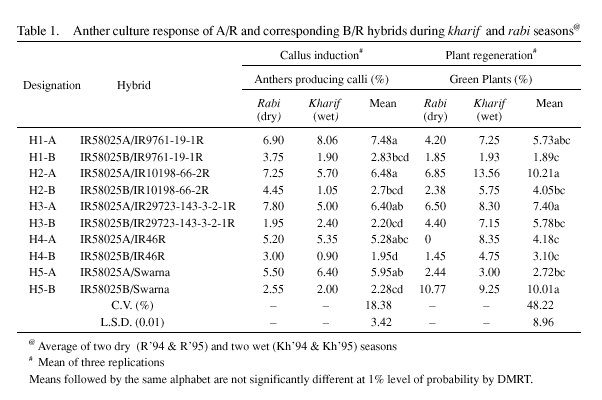
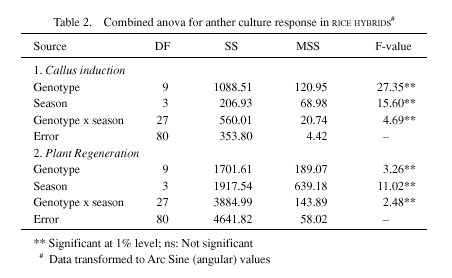
(B/R). Five sets of A/R hybrids involving a single cytoplasmic male sterile line, IR58025A and the five restorer lines: IR9761-19-1R, IR10198-66-2R, IR29723-143-3-2-1R, IR46R, and Swarna (hybrids designated as H1-A, H2-A, H3-A, H4-A, and H5-A); and the corresponding B/R hybrids with a single isonuclear male fertile line, IR58025B, crossed with the same set of five restorers (hybrids designated as H1-B, H2-B, H3-B, H4-B and H5-B), comprised the experimental materials (Table 1). Anthers from hybrid plants were plated for callus induction in N6 basic medium (Chu et al. 1978) supplemented with 2.0 mg/l, 2, 4-D and 0.5 mg/l, kinetin. For regeneration, MS basic medium (Murashige and Skoog 1962) with 2.5 mg/l BAP, 1.0 mg/l NAA and 0.5 mg/l kinetin was used. The experiment was repeated four times - two consecutive wet (kharif, 1994 & 1995) and dry (rabi, 1994 & 1995) seasons. The percent callus induction was relatively higher in all the five male sterile line (cms) based hybrids than the corresponding fertile B-line based hybrids in all the four seasons. Mean callusing frequency from the four experiments clearly showed that among the cms hybrids, H1-A topped with 7.48% callus induction, while H4-A responding the least with 5.28%. Comparatively, among the maintainer (B) line based hybrids, the highest callus induction frequency recorded was 2.83% in H1-B, and the lowest being in H4-B (1.95%). In all the four seasons similar trend, although frequency varied with different seasons, was observed for callus induction. Anther calli induced from both A/R and B/R hybrids were transferred to regeneration medium. Similar to callus induction, green plant regeneration frequency was also recorded higher among the A/R hybrids compared to the B/R hybrids. At least, in four of the five cms hybrids the green plant regeneration frequency was also recorded higher than their corresponding male fertile (B-line) hybrids. The data from all the four seasons were statistically analyzed. Separation of means by DMRT analysis confirmed the superior performance of A x R hybrid over B x R hybrids (Table1). An earlier study of anther culture of cms hybrids, conventional hybrids and normal varieties by Ling et al. (1978), clearly reported the better response of cms hybrids, in terms of average frequency of callus formation over the conventional hybrids and normal varieties. However, reports on anther culture response of cms lines in rice showed conflicting results. For instance, if the male sterility is caused by early microspore degeneration as in wild abortive (WA)
cytoplasm, a substantial decrease in callus production could occur due to reduced populations of potential microscopes that undergo androgenesis (Courtois and Tallebois 1990). Contrary to this, many workers found that callus induction in cms lines were rather high (Ling et al. 1978; Ge et al. 1985; Ling and Ma 2000). In the present study, we have not used the cms lines per se for anther culture. But the cms-based (A/R) hybrids in the background of WA cytoplasm and the corresponding main-tainer line based (B/R) hybrids were chosen to understand the phenomenon, on the assumption that the cms (A) line and the corresponding maintainer (B) line differ from each other on a cytoplasmic factor causing male sterility. Thus, our study while confirming the superior performance of the cms hybrids in anther culture also establishes the role of interaction of cytoplasmic and nuclear genomes in anther culturability. Most of the reports while recognizing the greater role of nuclear genome on callusing do not fail to underscore the significant influence of cytoplasm as well. Becraft and Taylor (1989) have brought out how changes in the level of interaction between nucleus and cytoplasm affect the anther culture response in wheat. Significant differences in the effect of cytoplasm on cell growth and development have been reported in different crops which include Brassica carinata, for shoot morphogenesis (Narasimhulu et al. 1989), maize, for callus growth (Nesticky et al. 1983) and somatic embryogenesis (Willman et al. 1989), barley for anther culture (Powel 1988) and potato for androgenetic competence (Singsit and Vielleux 1989). Observations in our study on the nature of gene action as well as the maternal effect governing the expression of the response to anther culture suggest the involvement of cytoplasmic and nuclear genes in anther culturability as earlier reported by Chu and Croughan (1989). Such an interaction is expected as the nucleus plays a regulatory role in the expression of the mitochondrial genome as well as the expression of nuclear genes encoding mitochondrial proteins. However, the underlying mechanism of nuclear-cytoplasmic interaction leading to higher response of anthers is still not clear. In our opinion, the difference between A/R and B/R may not only due to single/minor gene difference for male sterility but as well in the manner of expression of 'restored fertility' as in A/R and 'inherent fertility' in B/R. Further detailed studies like molecular mapping of anther culture responsive genes/alleles in rice chromosomes may be essential for better understanding of the whole mechanism. Acknowledgement The authors acknowledge the financial support from the Rockefeller Foundation, New York. They are also grateful to the Project Director, DRR, for providing the facilities. References Becraft, P.W., and Taylor, 1989. Effects of nucleus, cytoplasm and male sterile nucleus-cytoplasm combinations on callus initiation in anther culture of wheat. Euphytica 44: 235-240. Chu, C.C, Wang, C.C and C.S. Sun, 1978. The N6 medium and its application to anther culture of cereal crops. In: Proc. Symp. Plant Tissue Culture, Science Press, Peking, pp. 45-50. Chu, Q.R., and T.P. Croughan, 1989. Genetics of callus formation and plant regeneration in rice (Oryza sativa L.). In: Review of advances in plant biotechnology, 1985-1988. 2nd Int. Symp. Genetic Manipulation in Crops (Mujeeb-Kasi and L.A. Sitch Eds.), CIMMYT, Mexico and IRRI, Philippines, pp. 219-227. Courtois, B. and J. Tallebois, 1990. Anther culturability of rice lines bearing cytoplasmic male sterility. IRRN 15 (1): 7-8. Ge, M., Bai, H., and C. Tan, 1985. Anther culture of hybrid rice (indica x japonica and male sterile rice (wild-abortive)-Application in crop improvement. Phytobreedon 1 (2): 55-68. Ling, D. H., and Z.R. Ma, 2000. Restorable variation of cytoplasmic type of male sterile line in rice by in vitro culture. Chinese Science Bulletin. 45 (24): 2246-2250. Ling, T.H., Huang, H.S., Liang C.E., and P.Y. Chun, 1978. The study of anther culture in 'three lines' breeding and utilization of heterosis in Oryza sativa Subsp. hsien. Proc. Symp. Plant Tissue Cutlure, Science Press, Peking, pp. 213-222. Murashige, T., and F. Skoog, 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 15: 473-497. Narasimhulu, S.B., Chopra, V.L. and S. Prakash, 1989. The influence of cytoplasmic differences on shoot morphogenesis in Brassica carinata A. Euphytica 40: 241-243. Nesticky, M., Novak, F.J., A. Piovarci and M. Dolezelova, 1983. Genetic analysis of callus growth of maize (Zea mays L.) in vitro. Z. Pflanzenzuchtg. 91: 322-328. Powell, W., 1988. Diallel analysis of barley anther culture response. Genome 30: 152-157. Singsit, C. and R.E. Vielleux, 1989. Intra and inter specific transmission of androgenetic competence in diploid potato species. Euphytica 43: 105-107. Willman, M.R., S.M. Schroll and T.K. Hodges, 1989. Inheritance of somatic embryogenesis and plantlet regeneration from primary (Type I) callus in maize. In Vitro Cell. Deve. Biol. 25: 95-100. |
| RGN Home | Vol. 18>C. Research Notes> III. Genetics of physiological traits and others |
")
")
")
")
")
")
")
")

")





